

Cycle cellulaire et meiose
Tous les êtres vivants sont constitués d’unités invisibles à l’œil nu : les cellules.
Unité structurale et fonctionnelle du vivant.
Les organismes unicellulaires ne sont donc pas
directement accessibles à notre conscience, bien qu’ils représentent le plus grand nombre d’espèces présentes sur la Terre et constituent la plus grande partie de la biodiversité actuelle, tandis que les êtres multicellulaires, parfois de très grande taille, font partie de notre monde visible et familier.
Au plan structural, l’ensemble des êtres vivants est réparti en deux groupes :
Les procaryotes ou bactéries sont de très petite taille = 1µm ne présentant pas ou peu de compartimentation au sein de leur cytoplasme
Les eucaryotes sont des cellules de taille comprise entre 10-100µm, présentant un cytoplasme hautement structuré, contenant une grande diversité d’organites tels que : le noyau , les mitochondries, le réticulum endoplasmique……
Plan d’organisation procaryotique : Caractéristiques générales :
Sur le plan structural :
Organismes unicellulaires de forme sphérique ou en bâtonnets ;
Organismes plus complexes présentant une organisation filamenteuse (champignons) ;
Taille de 1µm à 10µm
A l’intérieur de la cellule pas d’organite, pas de compartimentation
Absence de cytosquelette et de processus endo- exocytose
Le patrimoine génétique n’est pas enfermé dans un noyau
Existence d’une double membrane limitante assurant un rôle de protection mécanique
La motilité : flagelle, par rotation, glissement
La division est simple, appelée la scissiparité , dans des conditions favorables de température l’Escherichia coli se multiplie toutes les 20 minutes.
Absence de sexualité vraie
Mode haploïde
Deux types les eubactèries et les archèes
Plan d’organisation eucaryotique : Sur le plan structural
Taille moyenne de 10 -100µm ( 1000 à un million de fois plus grosse que les procaryotes
Présence d’organites : mitochondries et les plastes (cellules végétales)
Présence de noyau renfermant le matériel génétique
Structures membranaires très développés : réticulum endoplasmique , appareil de Golgi
Vésicules de digestion intracellulaire : lysosomes, peroxysomes, endosomes.
Présence de vacuoles dans les cellules végétales
Présence de cytosquelette et présence de processus exocytose endocytose
Possèdent des chromosomes linéaires
Reproduction sexuée
Cellules animales et organismes animaux :
Exemple de cellule type : la cellule hépatique de taille de 20µm.
Les cellules animales possèdent en outre plusieurs caractéristiques liées au fait
qu’elles appartiennent à des organismes nécessairement pluricellulaires, où elles sont organisées en tissus, eux-mêmes assemblés en organes et appareils.
Au sein de certains tissus, les cellules sont étroitement accrochées entre elles par des dispositifs cytologiquement identifiables, appelés jonctions intercellulaires, dont la fonction première est d’assurer une cohésion mécanique.
Certaines permettent une communication directe entre les cytoplasmes et autorisent le passage de petites molécules d’une cellule à l’autre, alors que d’autres communiquent entre elles par l’intermédiaire de la matrice extracellulaire.
Les cellules animales présentent souvent des morphologies et des organisations internes complexes, en relation avec une fonction spécialisée ; cellules nerveuses (conduction du signal électrique), cellules musculaires (contraction) cellules rétinienne en bâtonnet (réception du signal lumineux).
Tableau I : comparaison structurale entre les cellules procaryotes et les cellules eucaryotes
| Procaryote | eucaryote | |
| noyau | absent | présent |
| Diamètre cellulaire | Petit 1 à10µm | Grand de 10 à100µm |
| Génome | Molécule ADN circulaire | Plusieurs molécules linéairesADN |
| ADN | Non associé avec les histones | Associé en complexe avecles histones |
| Quantité d’ADN | Assez faible | Assez grande |
| Organites liés à la membrane | absent | présent |
| cytosquelette | absent | présent |
La reproduction cellulaire :
Pour qu’une cellule se reproduise, 3 événements sont requis :
Information génétique doit être recopiée
Les copies d’information doivent être séparées
La cellule doit se diviser
Reproduction des cellules procaryotes :
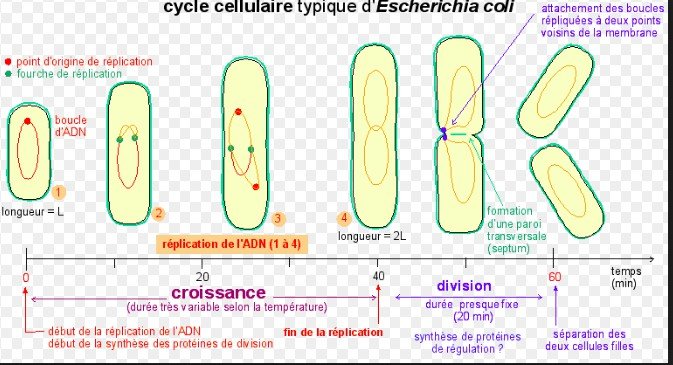
-la réplication du chromosome circulaire commence par un site appelé origine de réplication.
les deux origines nouvellement répliquées s’éloignent l’une de l’autre en direction des extrémités de la cellule.
Chez certaines bactéries au moins, des protéines se lient à proximité de l’origine de réplication et attachent les nouveaux chromosomes à la membrane plasmique aux extrémités opposées (« pôles ») de la cellule.
une nouvelle paroi cellulaire se forme entre les chromosomes, produisant deux cellules contenant chacune un exemplaire identique du chromosome.
Reproduction des cellules eucaryotes :
Réplication de l’ADN, séparation des copies et division du cytoplasme.
chromosomes eucaryotiques : chaque espèce à un nombre caractéristique de chromosome : 48 pour la pomme de terre, 8 pour les drosophiles, 46 pour l’être humain.
La majorité des eucaryotes ont deux jeux de chromosomes, ceci est la conséquence de la reproduction sexuée.
Un hérité du père et l’autre de la mère, l’ensemble forme une paire d’homologues.
Les cellules humaines contiennent 23 paires d’homologues.
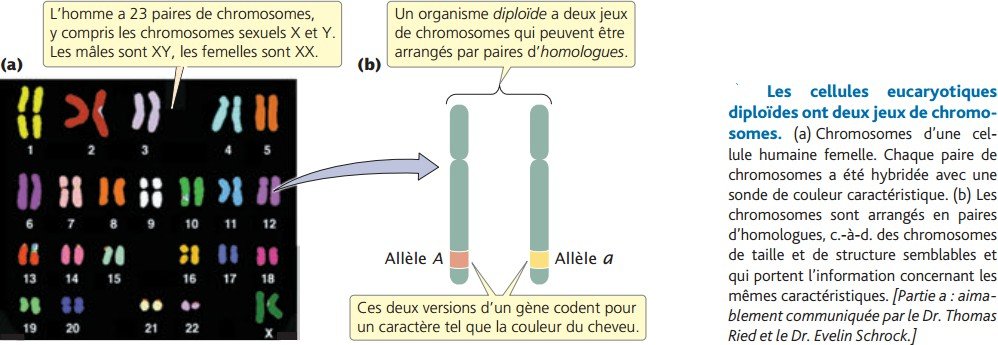
Les deux chromosomes d’une paire d’homologues sont généralement de taille similaire et
chacun contient l’information génétique pour le même jeu de caractère héréditaire .Si un gène d’un chromosome particulier code pour un caractère héréditaire tel que la couleur des cheveux, une autre copie du gène (chaque copie étant appelée un allèle), à la même position sur le chromosome homologue, code aussi pour la couleur.
Structure du chromosome :
La structure d’un chromosome eucaryote est bien plus grande et plus complexe que celui d’un procaryote.
Chaque chromosome à l’état non répliqué ne contient qu’une seule molécule d’ADN.
Les molécules d’ADN sont linéaires, hautement repliées et condensées. ; Sous forme étirées elles mesurent plusieurs centimètres soit mille fois qu’un noyau.
Ainsi pour être contenu dans un petit volume de noyau, chaque molécule d’ADN
s’enroule de façon serrée autour de molécules d’histone pour former des bâtonnets de chromosomes.
Les molécules de chromosomes sont observées avant la division cellulaire.
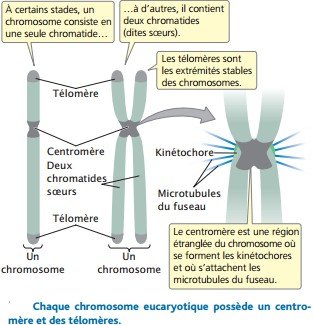
Un chromosome fonctionnel possède trois éléments essentiels : un centromère, une paire de télomère, des origines de réplications
Le centromère est le point d’attachement des microtubules du fuseau qui sont les filaments responsables du mouvement des chromosomes à la division cellulaire.
Avant la division cellulaire, un complexe de protéines appelé kinétochore s’assemble sur le centromère.
Ensuite, les microtubules du fuseau s’attachent au kinétochore.
Les télomères sont les extrémités naturelles d’un chromosome linéaire.
Les télomères confèrent une stabilité au chromosome.
Les origines de réplication : sont les sites où commence la synthèse d’ADN ; En préparation à la division cellulaire, chaque chromosome se réplique
Les deux copies initialement identiques, appelées chromatides sœurs, sont maintenues ensemble par le centromère.
Chaque chromatide sœur contient une seule molécule d’ADN.
Cycle cellulaire:
Définition : le cycle cellulaire est l’histoire de la vie d’une cellule, les stades par lesquels elle passe d’une division à la suivante.
Le capital génétique est transmis des parents aux cellules filles.
Un nouveau cycle commence quand une cellule s’est divisée et a produit deux nouvelles cellules.
Chaque nouvelle cellule métabolise, croît et se développe. À la fin de son
cycle, elle se divise et produit deux cellules qui peuvent alors suivre des cycles cellulaires supplémentaires.
La progression dans le cycle cellulaire est contrôlée à des points clés appelés points de contrôle.
Les différentes phases du cycle cellulaire :
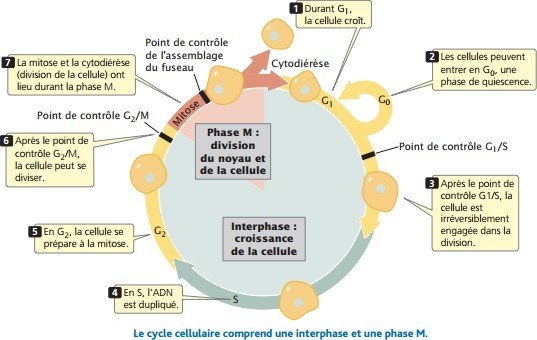
Le cycle cellulaire comporte deux phases une interphase et la phase M
La première phase interphase : la période entre les divisions cellulaires pendant laquelle la cellule croît, se développe, et se prépare à la division.
La deuxième phase est la phase M ( mitotique ) correspond à la division active.
La phase M comprend la mitose, le processus de division du noyau, et la cytodiérèse ou division cytoplasmique.
L’interphase :
L’interphase est la longue période de croissance et de développement entre deux divisions cellulaires.
L’interphase comporte plusieurs points de contrôle qui
régulent le cycle cellulaire.
Les points de contrôle sont nécessaires pour empêcher la prolifération de cellules contenant des chromosomes abîmés ou auxquelles manquent des chromosomes.
L’interphase est subdivisée en trois phases : G1, S, et G2
Phase G1 ( gap1 : intervalle): la cellule croît et synthétise les protéines nécessaires à sa division ; ce stade dure généralement plusieurs heures.
Le point de contrôle G1/S, juste avant la fin de G1.
Ce point de contrôle maintient la cellule en G1 jusqu’à ce qu’elle possède toutes les enzymes nécessaires à la réplication de l’ADN.
Une fois franchi ce point de contrôle, la cellule est irréversiblement engagée dans la division.
Les cellules peuvent sortir du cycle cellulaire actif en réponse à des signaux régulateurs et passer dans une phase de quiescence appeléeG0, qui correspond à un état stable, et pendant laquelle les cellules ne changent pas de taille.
Elles peuvent rester en G0 pendant des temps variables, voire indéfiniment, ou regagner G1 et reprendre le cycle actif.
De nombreuses cellules n’entrent jamais en G0 et effectuent continuellement des cycles
Phase S : après la phase G1, la cellule rentre dans la phase de synthèse de l’ADN, chaque chromosome est dupliqué.
Avant la phase S, chaque chromosome est constitué d’une chromatide, après la phase S chaque chromosome contient deux chromatides.
PhaseG2 : dans cette phase se produisent plusieurs autres événements biochimiques nécessaires à la division cellulaire.
Le point de contrôle G2/M est atteint avant la phase G2.
Une fois franchi, la cellule entre dans la phase M.
La durée de l’interphase varie selon le type cellulaire, Une cellule de mammifère typique en cours de division passe environ 10 heures en G1, 9 heures en S, et 4 heures en G2.
Pendant toute l’interphase, les chromosomes sont dans un état relâché, mais pas désenroulé, et ils ne sont pas visibles au microscope optique.
Mitose :
La phase M est la période du cycle cellulaire pendant laquelle les chromosomes (en fait, les chromatides sœurs) se séparent et la cellule se divise.
La phase M est subdivisée en 6 stades : 5 stades (prophase,prométaphase, métaphase, anaphase et télophase) ET cytodièrèse
Prophase : les chromosomes deviennent visibles au microscope durant cette phase.
Le fuseau mitotique se forme à partir d’une paire de centrosomes qui migrent vers les extrémités opposées de la cellule. À l’intérieur de chaque centrosome se trouve un organite spécialisé, le centriole, qui est aussi composé de microtubules.
Prométaphase : durant cette phase, dégradation de la membrane nucléaire.
Les microtubules du fuseau qui, jusqu’à ce point, étaient à l’extérieur du noyau, entrent dans la région nucléaire.
Pour chaque chromosome, un microtubule d’un des centrosomes s’attache au kinétochore d’une des chromatides sœurs tandis qu’un microtubule du centrosome opposé s’attache à l’autre chromatide sœur, de sorte que chaque chromosome se trouve relié aux deux centrosomes.
Les microtubules s’allongent et se raccourcissent, poussant et tirant les chromosomes.
Métaphase : les chromosomes s’alignent dans un plan central, le plan équatorial, à équidistance des deux centrosomes.
Anaphase : commence quand les chromatides sœurs se séparent et migrent vers les pôles opposés du fuseau.
Les chromatides séparées sont considérées comme des chromosomes à part entière.
Télophase : les chromosomes arrivent à proximité des pôles du fuseau.
Une membrane nucléaire se reforme autour de chaque jeu de chromosomes, créant deux noyaux distincts dans la cellule.
Les chromosomes se décondensent et s’allongent, reprenant un aspect diffus non visible au microscope.
Dans de nombreux types cellulaires, la division du cytoplasme (cytodiérèse) se produit simultanément à la télophase.
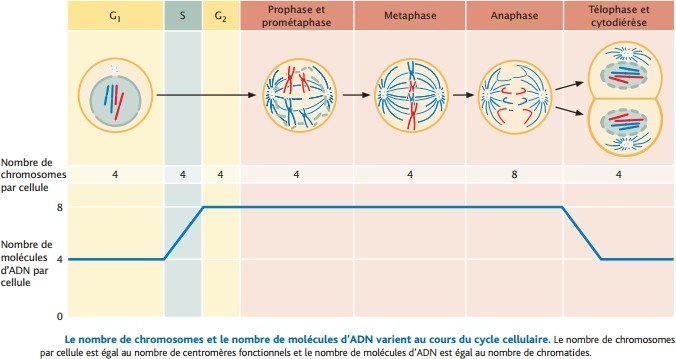
Le nombre de chromosome et de molécules d’ADN :
-Au début de G1, cette cellule diploïde a deux jeux complets de chromosomes, hérités de la cellule dont elle est issue.
Chaque chromosome consiste en une seule chromatide – une seule molécule d’ADN – et donc, il y a quatre molécules d’ADN en G1.
– Pendant la phase S, chaque molécule d’ADN est copiée. le nombre de chromosomes est inchangé parce que les chromatides sœurs sont attachées et partagent un seul centromère fonctionnel.
Quatre chromosomes comprenant chacun deux chromatides, et donc, elle contient huit molécules d’ADN.
Pendant la prophase, la prométaphase, et la métaphase, la cellule contient quatre chromosomes et huit molécules d’ADN.
À l’anaphase, les chromatides sœurs se séparent.
Chacune a désormais son propre centromère et est donc considérée comme un chromosome à part entière. La cellule contient huit chromosomes, consistant chacun en une seule chromatide, et elle contient toujours huit molécules d’ADN.
Après la cytodiérèse, les huit chromosomes (les huit molécules d’ADN) sont répartis également entre deux cellules, de sorte que chaque nouvelle cellule contient quatre chromosomes et quatre molécules d’ADN, soit le même nombre qu’au début du cycle cellulaire.
Les points de contrôle du cycle cellulaire
Les divisions cellulaires sont essentielles au développement des organismes eucaryotes pluricellulaires, mais aussi à leur survie lorsqu’ils ont atteint l’âge adulte (notion d’homéostasie générale).
Chez les eucaryotes, le cycle cellulaire est contrôlé par un système mettant en jeu de nombreuses protéines régulatrices très conservées au cours de l’évolution.
Des dérèglements de ce système de contrôle peuvent entraîner une prolifération cellulaire excessive et un cancer.
Il existe trois points de contrôle de régulation du cycle cellulaire :
– Point de contrôle en G1 ou point de restriction R :
L’environnement est-il favorable ?
Des signaux de croissance et de division sont-ils présents dans le milieu ? L’ADN est-il endommagé ?
La cellule poursuit la division cellulaire que si les conditions extérieures sont favorables (facteurs de croissance)
Point de contrôle en G2 : Tout l’ADN est-il répliqué ? L’ADN est-il endommagé ?
L’entrée en M n’est possible que si l’ADN est totalement répliqué et réparé
Point de contrôle en M :
Tous les chromosomes sont-ils attachés de façon bipolaire ?
Les protéines de point de contrôle :
La cellule dispose de systèmes de régulation hautement perfectionnés.
Dans le premier cas, (régulation du cycle), ce sont essentiellement des kinases cycline-dépendantes, les Cdk, qui interviennent.
Dans le second cas, d’autres molécules interviennent dans différents mécanismes de surveillance du cycle pour inhiber les Cdk de la régulation du cycle et arrêter le cycle.
Les protéines Cdk : Les kinases cycline-dépendantes (Cdk) sont une famille de
protéineskinases régulant la progression dans le cycle cellulaire.
Chaque Cdk reconnaît spécifiquement un substrat (ou un groupe de substrats) qu’elle phosphoryle en utilisant le phosphate de l’ATP.
Les Cdk forment des complexes hétérodimériques avec les cyclines, leurs sous unités régulatrices.
Le cycle cellulaire est contrôlé par au moins 6 complexes Cycline / Cdk différents qui interviennent à des moments précis du cycle cellulaire.
Tableau : Les complexes cycline-Cdk et leur rôle au cours du cycle cellulaire

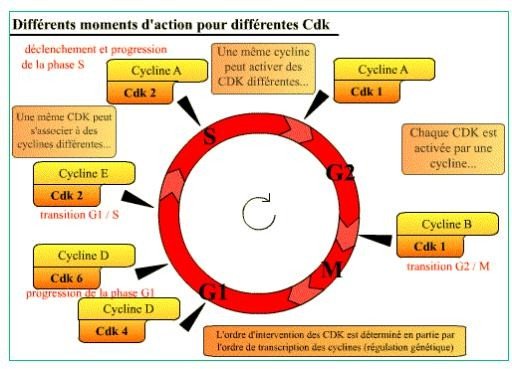
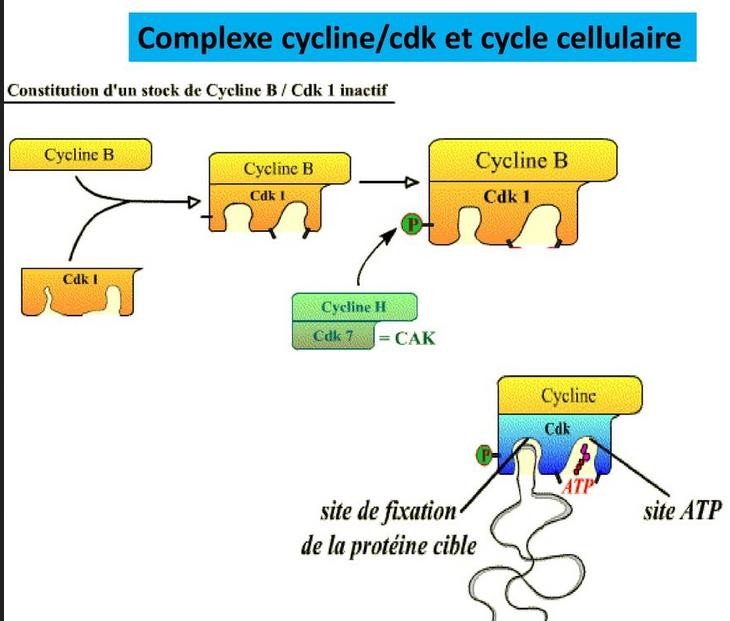
Figure : complexes Cdk-cycline et cycle cellulaire
Les protéines activatrices et inhibitrices CdK :
Les Cdk sont activées, par :
des phosphatases : Cdc 25 [Déphosphorylations « activatrices »] ;
des kinases : CAK (« Cdk Activating Kinase » [Phosphorylations « activatrices »].
Les Cdk sont inhibées par :
des protéines inhibitrices (inhibiteurs physiologiques), les CKI (Cdk Inhibitor) : p16, p21, p27, qui agissent sur les complexes Cycline / Cdk ;
une kinase : Wee 1 [responsable de Phosphorylations “inhibitrices”] qui agit sur la Cdk1 en la phosphorylant
La protéine P53 : p53 est un facteur de transcription qui active la transcription de nombreux gènes, dont celui qui code p21 CKI.P53 protéine de contrôle de l’intégrité de l’ADN.
En l’absence de lésion de l’ADN, la P53 est faible.
En présence de lésion de l’ADN, augmentation de la concentration de la P53 et par conséquent activation de la P21 CdK inhibitrice bloquant l’entrée en phase S
la reproduction sexuée par le processus de méiose :
La reproduction sexuée comporte deux processus.
Le premier est la méiose qui produit des gamètes dans lesquels le nombre de chromosomes est réduit de moitié.
Le second est la fécondation, dans laquelle deux gamètes haploïdes fusionnent pour rétablir le nombre de chromosomes caractéristique de l’état diploïde original.
La méiose : la méiose est précédée d’une interphase qui comprend des stades G1, S et G2.
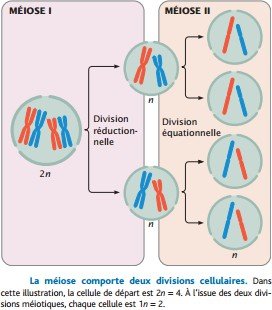
La méiose comprend deux processus distincts, la méiose I et la méiose II, qui comprennent chacun une division cellulaire.
La première division, à la fin de la méiose I, est appelée division réductionnelle parce qu’elle réduit de moitié le nombre de chromosomes.
La seconde division, à la fin de la méiose II, est parfois appelée division équationnelle.
Les stades de la méiose II sont similaires à ceux de la mitose.
Toutefois, la méiose II diffère de la mitose en ce sens que le nombre de chromosomes a déjà été réduit de moitié pendant la méiose I.
Méiose I :
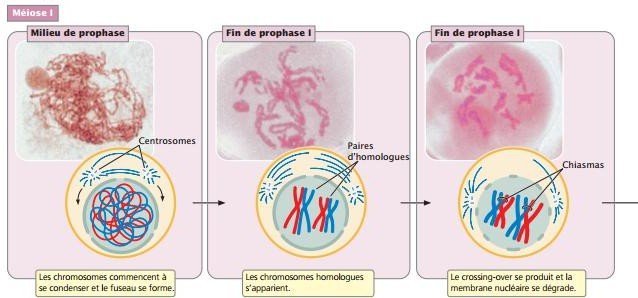
La prophase est un stade long pendant lequel les chromosomes homologues s’apparient et des crossing-over se produisent, au cours desquels les chromosomes échangent de l’information génétique.
Les centromères des chromosomes appariés s’écartent et les homologues restent associés par des chiasmas qui sont la manifestation visible des crossing- over ; À la fin de la prophase I, la membrane nucléaire disparaît et le fuseau se forme.
La métaphase I débute lorsque les paires de chromosomes homologues s’alignent le long du plan équatorial.
L’anaphase I est marquée par la séparation des chromosomes homologues.
Les deux chromosomes d’une paire sont attirés vers les pôles opposés.
A la télophase I les chromosomes arrivent aux pôles du fuseau et le cytoplasme se divise.
L’intervalle entre la meiose I et la meiose II est l’interkinèse.
Une membrane nucléaire se reconstitue autour des chromosomes rassemblés à chaque pôle, le fuseau se dégrade et les chromosomes se relâchent.
Les cellules entrent alors en prophase II, où les événements de l’interkinèse sont inversés : les chromosomes se recondensent, le fuseau se reforme, et l’enveloppe nucléaire se dégrade à nouveau.
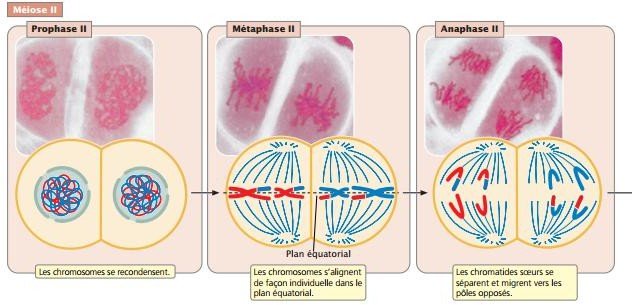
Méiose II
Ces cellules passent directement de la cytodiérèse à la métaphase II qui est semblable à la métaphase mitotique : les chromosomes individuels s’alignent dans le plan équatorial.
À l’anaphase II, les kinétochores des chromatides sœurs se séparent et les chromatides sont attirées vers les pôles opposés.
Chaque chromatide est désormais un chromosome à part entière.
À la télophase II, les chromosomes arrivent aux pôles du fuseau, une enveloppe nucléaire se reforme autour d’eux, et le cytoplasme se divise.
La méiose, source de variation génétique
À part les vrais jumeaux, deux individus issus de mêmes parents ne sont pas génétiquement identiques.
Cela est dû au fait que, longtemps avant la fusion des deux gamètes, deux types de brassage génétique ont déjà eu lieu au cours de la méiose.
Brassage inter chromosomique : assortiment indépendant des chromosomes
Le premier type de brassage résulte de la répartition au hasard des homologues maternels et paternels entre les cellules filles au cours de la méiose I : chaque gamète reçoit un mélange différent de chromosomes maternels et paternels.
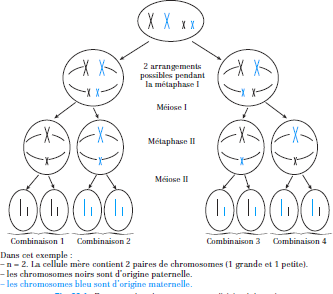
Figure : brassage des chromosomes en division 1 de meiose Crossing-over (brassage intra-chromosomique)
Le crossing-over (ou enjambement) est un processus qui a lieu au coursde la prophase I et dans lequel des segments de chromosomes homologues sont échangés.
Le crossing-over implique la cassure des doubles hélices d’ADN maternelle et paternelle de chacune des deux chromatides et leur échange de fragments de façon réciproque par un processus appelé recombinaison génétique.
Les événements de recombinaison génétique sont catalysés par les nodules de recombinaison.
Ce sont des complexes protéiques multienzymatiques XM
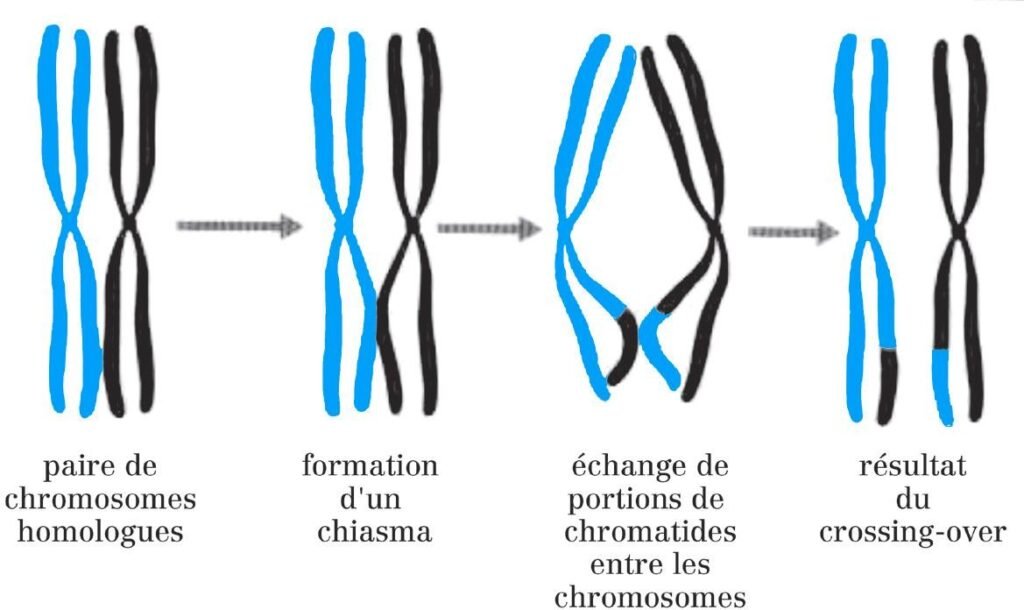
Figure : Crossing-over lors de la prophase I de méiose
La liaison génétique ou linkage : Lorsque deux gènes sont situés proches l’un de l’autre sur le même chromosome, leur assortiment n’est pas indépendant.
La liaison de deux gènes sur le même chromosome est appelé liaison génétique (linkage en anglais).
La fréquence de la recombinaison qui est la proportion des gamètes recombinés est utilisée pour exprimer et estimer la distance entre deux gènes.
Un pourcentage de liaison est égal à un centiMorgan (cM).
Si la fréquence en gamètes recombinés est égale à la fréquence en gamètes parentaux donc égale à 1/2, on dit que les deux gènes sont non liés ou indépendants (génétiquement) Si la fréquence de recombinaison est inférieure à la fréquence en gamètes parentaux, alors il y a une liaison génétique.