Biochimie · Enzymologie · Pharmacologie moléculaire · Mis à jour 2025
Introduction : Pourquoi les enzymes allostériques sont-elles essentielles ?
Le modèle de Michaelis-Menten a profondément marqué la biochimie enzymatique. Pourtant, il existe tout un groupe d’enzymes qui lui échappent : les enzymes allostériques — de véritables interrupteurs moléculaires au cœur du métabolisme cellulaire.
Contrairement aux enzymes classiques, ces protéines ne se contentent pas de catalyser des réactions. Elles intègrent des signaux provenant de l’environnement cellulaire et ajustent leur activité en temps réel, assurant une régulation fine et précise des voies métaboliques.
Sur le plan médical, les enzymes allostériques représentent des cibles thérapeutiques de premier plan. Les modulateurs allostériques offrent une spécificité accrue, des effets secondaires réduits et des possibilités d’action synergique — là où les médicaments classiques montrent leurs limites.
“Comprendre les enzymes allostériques, c’est comprendre comment la cellule régule son propre métabolisme — et comment la médecine peut exploiter ces mécanismes pour traiter de nombreuses maladies.”
I. Généralités sur les Enzymes Allostériques
1. Définition de l’allostérie
Le terme allostérie vient du grec allos (autre) et stéros (espace, site) : il désigne littéralement un « autre site que le site catalytique ». Ce concept a été introduit par Monod, Changeux et Jacob en 1963 lors de leurs travaux fondateurs sur la régulation enzymatique.
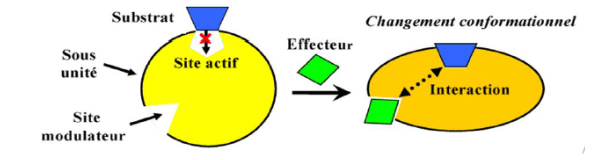
L’allostérie désigne la variation de la conformation spatiale d’une protéine suite à la liaison réversible d’un composé hors du site catalytique, entraînant une modification — activatrice ou inhibitrice — de son activité. Ce processus est exclusif aux protéines de structure quaternaire.
💡 À noter — L’allostérie ne concerne pas seulement les enzymes : elle régit aussi les récepteurs membranaires, les protéines de transport comme l’hémoglobine, et les facteurs de transcription.
2. Caractéristiques des enzymes allostériques
Une enzyme allostérique se distingue par un ensemble de propriétés structurales et cinétiques :
Structure quaternaire obligatoire — au moins 2 sous-unités (protomères)
Sites multiples — un ou plusieurs sites catalytiques + un ou plusieurs sites allostériques distincts
Liaison réversible et non covalente des effecteurs sur leurs sites spécifiques
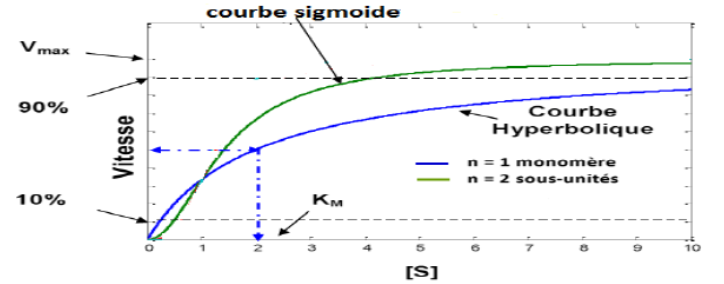
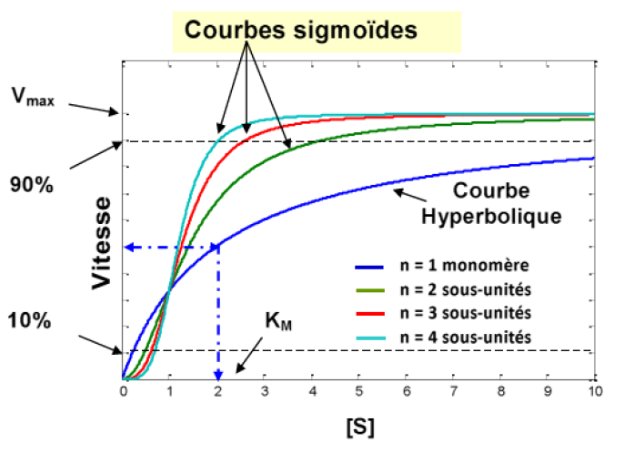
Courbes cinétiques sigmoïdes (en forme de S), et non les courbes hyperboliques de Michaelis-Menten
Grande sensibilité aux variations de concentration en substrat dans une gamme intermédiaire
Rôle clé dans la régulation métabolique — contrôle des voies énergétiques et biosynthétiques
Fait important : lorsqu’une enzyme allostérique est dénaturée (perte de la structure quaternaire), elle perd ses propriétés allostériques et adopte un comportement purement michaélien. Cela confirme que la structure quaternaire est indispensable à la régulation allostérique.
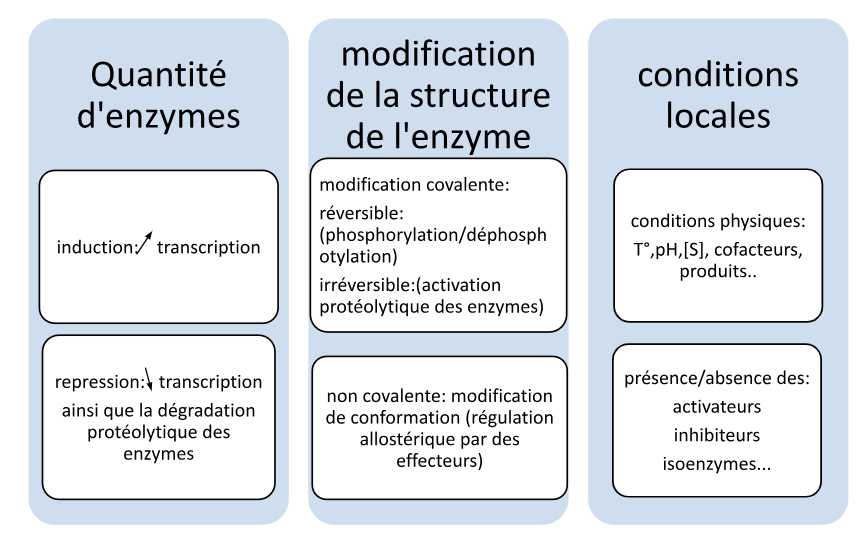
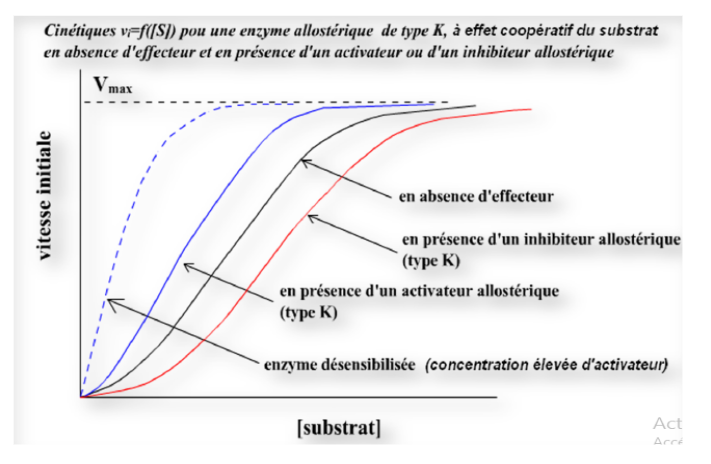
3. Classification : systèmes K, V et mixtes
Type de système
Paramètre modifié
Description
Système K
Km uniquement
L’effecteur modifie l’affinité de l’enzyme pour son substrat — la Vmax reste inchangée
Système V
Vmax uniquement
L’effecteur modifie la vitesse maximale — le Km reste constant
Système mixte
Km + Vmax
L’effecteur modifie à la fois l’affinité et la vitesse maximale
4. La transition allostérique : formes T et R
Les enzymes allostériques existent en équilibre entre deux états conformationnels :
Forme T (Tendue) — conformation compacte à faible affinité pour le substrat. État adopté en l’absence de substrat ou en présence d’inhibiteurs.
Forme R (Relâchée) — conformation plus ouverte à forte affinitépour le substrat. État favorisé par la liaison du substrat ou d’activateurs allostériques.
Une transition allostérique correspond au passage T → R (augmentation d’affinité) ou R → T (diminution d’affinité), déclenché par le substrat ou un effecteur hétérotrope.
II. La Modulation Allostérique
1. La coopérativité : le principe fondamental
La coopérativité est la propriété clé qui rend les enzymes allostériques si efficaces comme régulateurs. Elle désigne le fait que la liaison d’un substrat à une sous-unité influence l’affinité des autres sous-unités pour ce même substrat.
Concrètement : quand une sous-unité change de conformation suite à la liaison d’un substrat, elle « communique » ce changement à ses voisines, qui adoptent à leur tour une conformation plus favorable. Ce phénomène inter-sous-unités engendre la courbe sigmoïde caractéristique.
2. Effets homotropes
L’effet homotrope se produit lorsque le substrat lui-même joue le rôle de modulateur en se fixant à la fois sur le site catalytique et sur le site allostérique.
Homotrope positif (T → R) : la fixation d’une molécule de substrat augmente l’affinité pour les suivantes. « L’appétit vient en mangeant. »
Homotrope négatif (R → T) : la fixation d’une molécule de substrat réduit l’affinité pour les suivantes. Phénomène plus rare. « La soif s’en va en buvant. »
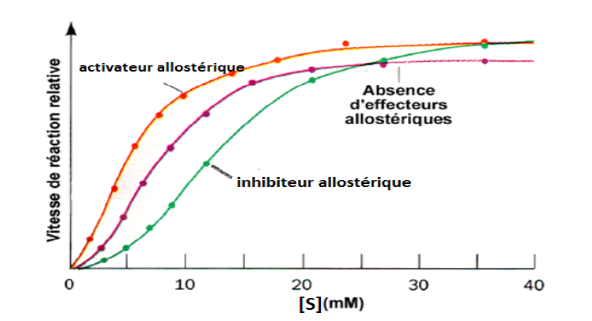
3. Effets hétérotropes
L’effet hétérotrope implique des molécules structuralement différentes du substrat — les effecteurs allostériques — qui modifient l’affinité de l’enzyme sans en être elles-mêmes des substrats.
Activateurs allostériques (effecteurs positifs) : favorisent la transition T → R, augmentant l’affinité pour le substrat.
Inhibiteurs allostériques (effecteurs négatifs) : maintiennent ou favorisent la conformation T, réduisant l’activité enzymatique.
L’effet hétérotrope est capital car il permet la communication entre voies métaboliques différentes : un métabolite d’une voie peut réguler une enzyme d’une autre voie, assurant une coordination globale du métabolisme cellulaire.
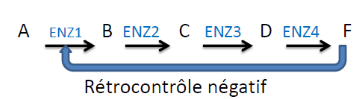
4. La rétroinhibition (feedback inhibition)
La rétroinhibition est l’un des mécanismes de régulation métabolique les plus élégants du vivant. Son principe : le produit final d’une voie métabolique inhibe allostériquement l’enzyme qui catalyse la première réaction de cette voie.
Résultat : dès que le produit final s’accumule en excès, il bloque automatiquement sa propre production — évitant tout gaspillage énergétique. Quand sa concentration baisse, l’inhibition se lève et la production reprend.
🧬 Application universelle — La rétroinhibition régule des voies fondamentales : biosynthèse des acides aminés, synthèse des nucléotides, glycolyse. Ce mécanisme est conservé de la bactérie à l’humain — preuve de son importance évolutive.
III. Cinétique Allostérique
1. La courbe sigmoïde : signature diagnostique
La courbe sigmoïde (en S), obtenue en traçant Vo en fonction de [S], est le critère diagnostique fondamental d’une enzyme allostérique. Elle contraste fortement avec la courbe hyperbolique des enzymes michaéliennes.
Cette sigmoïde peut être interprétée comme la superposition de deux courbes hyperboliques : l’une correspondant à l’état T (faible affinité) et l’autre à l’état R (forte affinité). Elle confère à l’enzyme une sensibilité remarquable dans une gamme intermédiaire de concentration en substrat — comportement d’un véritable interrupteur moléculaire.
2. L’équation de Hill (1910)
En 1910, A.V. Hill proposa empiriquement la première équation pour décrire la liaison coopérative. Transposée à la cinétique enzymatique :
V₀ = Vmax × [S]ʰ / (K₀.₅ʰ + [S]ʰ)
Le coefficient de Hill (h) quantifie le degré de coopérativité :
h = 1 → pas de coopérativité (comportement michaélien)
Les sous-unités (protomères) sont toutes équivalentes ; l’oligomère possède au moins un axe de symétrie.
Chaque protomère existe dans deux états : R (relâché, forte affinité) et T (tendu, faible affinité), en équilibre même sans ligand.
Les deux états ont des affinités différentes pour un ligand donné.
Toute transition conformationnelle conserve la symétrie de l’oligomère : le changement est concerté (toutes les sous-unités simultanément).
Conséquence directe : pas d’espèces hybrides dans le même oligomère. Le modèle MWC est un modèle « tout ou rien ». Limitation : il ne peut expliquer que la coopérativité positive.
Fondé sur le concept d’ajustement induit, ce modèle propose :
En l’absence de ligand, la protéine existe dans un seul état conformationnel — pas d’équilibre T ⇌ R préexistant.
La liaison d’un ligand induit un changement séquentiel dans la sous-unité liée, puis transmis aux voisines par contacts inter-protomériques.
Ce modèle explique à la fois la coopérativité positive et négative.
Critère
Modèle MWC
Modèle KNF
Changement conformationnel
Concerté
Séquentiel
État initial
Équilibre T ⇌ R préexistant
Un seul état
Espèces hybrides
Impossibles
Possibles
Type de coopérativité
Positive seulement
Positive et négative
Concept clé
Symétrie conservée
Ajustement induit
IV. Applications Médicales et Perspectives Thérapeutiques
1. Les enzymes allostériques comme cibles médicamenteuses
L’allostérie est aujourd’hui au cœur d’une révolution pharmacologique. Les modulateurs allostériques présentent des avantages décisifs par rapport aux médicaments orthostériques classiques :
Spécificité accrue — les sites allostériques sont souvent moins conservés entre membres d’une même famille protéique
Effets secondaires réduits — modulation de l’activité plutôt que blocage total, préservant une activité résiduelle physiologique
Synergies possibles — action conjointe avec des médicaments orthostériques pour une efficacité combinée
Cibles “non-druggables” — certaines protéines sans site actif défini peuvent être ciblées via leurs sites allostériques
2. Médicaments allostériques approuvés
Médicament
Cible allostérique
Indication thérapeutique
Benzodiazépines (1960s)
Récepteur GABA-A
Anxiété, épilepsie — parmi les premiers médicaments allostériques
Maraviroc (2007)
Récepteur CCR5
Infection VIH — modulateur allostérique négatif
Cobimetinib (2015)
MEK1/2
Mélanome métastatique (mutation BRAF)
Adagrasib (MRTX-849)
KRAS G12C (site S-IIP)
Cancer du poumon — >1000× plus sélectif que le KRAS sauvage
3. L’IA et la dynamique moléculaire au service de l’allostérie
Depuis 2020, l’intelligence artificielle transforme la découverte des sites allostériques. Des outils comme AlphaFold2, PASSer et AlloReverse permettent de prédire les sites allostériques à partir de la seule séquence protéique.
Les simulations de dynamique moléculaire (MD), couplées à l’apprentissage automatique, permettent d’identifier des poches cryptiques — sites allostériques transitoires invisibles dans les structures cristallographiques statiques, mais présents lors des mouvements conformationnels naturels de la protéine.
🚀 Perspective 2025 — Les anticorps allostériques constituent une classe émergente de thérapeutiques. Contrairement aux petites molécules, ils offrent une sélectivité inégalée pour cibler des récepteurs membranaires (GPCRs, canaux ioniques) longtemps considérés comme non-ciblables.
Conclusion : L’allostérie, un langage universel du vivant
Les enzymes allostériques sont bien plus que de simples catalyseurs. Ce sont des capteurs moléculaires sophistiqués qui permettent à la cellule de lire son environnement et d’adapter son métabolisme en temps réel.
Du modèle empirique de Hill (1910) aux outils d’intelligence artificielle d’aujourd’hui, la recherche sur l’allostérie a parcouru un chemin extraordinaire. Ce qui était un phénomène cinétique curieux est devenu l’un des mécanismes biologiques les mieux exploités en pharmacologie moderne.
Comprendre les enzymes allostériques, c’est comprendre à la fois comment la vie se régule elle-même, et comment la médecine peut intervenir avec précision pour corriger les dérèglements à l’origine des maladies.
Points clés à retenir
Les enzymes allostériques possèdent une structure quaternaire et des sites allostériques distincts du site catalytique
Leur cinétique est sigmoïde, reflétant la coopérativité entre sous-unités protomériques
Deux modèles théoriques principaux : le modèle concerté MWC et le modèle séquentiel KNF
La rétroinhibition est un mécanisme de régulation métabolique universel, conservé de la bactérie à l’humain
Les modulateurs allostériques constituent une classe de médicaments à fort potentiel — spécificité accrue, effets secondaires réduits
L’IA et la dynamique moléculaire révolutionnent la découverte de nouveaux sites allostériques cryptiques
L’anesthésie est l’une des avancées médicales les plus révolutionnaires de l’histoire. Sans elle, pas de chirurgie sans douleur, pas d’interventions...