Les enzymes sont les moteurs moléculaires du vivant. Elles accélèrent les réactions chimiques des milliers, voire des millions de fois par rapport à ce qui se passerait spontanément. Mais comment mesure-t-on leur efficacité ? Comment comprendre à quelle vitesse elles travaillent, et quels facteurs influencent leur activité ? C’est précisément l’objet de la cinétique enzymatique.
Dans cette première partie, nous posons les bases essentielles : définitions, principes mathématiques, modèle de Michaelis-Menten et interprétation des constantes cinétiques. Un savoir fondamental pour tout étudiant en biochimie, pharmacologie ou médecine.
I. Qu’est-ce que la Cinétique Enzymatique ?
1.1 Définition
La cinétique enzymatique est la branche de la biochimie qui étudie la vitesse des réactions chimiques catalysées par des enzymes, ainsi que les facteurs qui l’influencent. Elle permet concrètement de :
Calculer la vitesse catalytique maximale d’une enzyme (Vmax)
Déterminer les affinités de liaison des substrats et des inhibiteurs pour l’enzyme
Évaluer l’influence de paramètres extérieurs tels que le pH, la température ou la présence d’effecteurs
En pratique, la cinétique enzymatique est un outil indispensable en pharmacologie (conception de médicaments), en diagnostic médical (dosage d’enzymes sériques) et en biotechnologie industrielle (optimisation de biocatalyseurs).
1.2 Les Différentes Phases d’une Réaction Enzymatique
Lorsqu’on suit l’évolution d’une réaction enzymatique dans le temps, on distingue trois phases :
Phase initiale (ordre 0 apparent) : la concentration en produit [P] augmente de manière linéaire avec le temps. C’est dans cette fenêtre que l’on mesure la vitesse initiale V₀.
Phase intermédiaire : la vitesse ralentit progressivement à mesure que le substrat s’épuise ou que les produits s’accumulent.
Phase d’équilibre : [P] atteint un plateau. La vitesse nette est nulle, l’équilibre thermodynamique est établi.
La mesure se fait toujours en vitesse initiale — c’est-à-dire au tout début de la réaction — pour éviter l’accumulation de produit et garantir des conditions reproductibles.
II. Principes de Base : Vitesse, Molécularité et Ordre d’une Réaction
2.1 La Vitesse d’une Réaction
Pour une réaction simple A → P, la vitesse est définie comme la variation de concentration d’un réactif ou d’un produit par unité de temps :
V = -d[A]/dt = +d[P]/dt = K · [A]
Où K est la constante de vitesse (ou constante cinétique), qui dépend de la température et de la nature de la réaction.
2.2 Molécularité d’une Réaction
La molécularité désigne le nombre de molécules qui participent simultanément à l’étape élémentaire d’une réaction :
Mono-moléculaire : A → P (ex. : isomérisation)
Bi-moléculaire : A + B → P (ex. : la plupart des réactions enzymatiques)
Tri-moléculaire : rare, car la probabilité que trois molécules se rencontrent simultanément est très faible
2.3 Ordre d’une Réaction
Pour une réaction générale αA + βB → P + Q, la loi de vitesse s’écrit :
V = K · [A]^α · [B]^β
L’ordre global n = α + β est la somme des exposants des concentrations dans l’expression de la vitesse.
Ordre
Expression de la vitesse
Signification
Ordre 0
V = K
Vitesse constante, indépendante de [S]
Ordre 1
V = K · [A]
Vitesse proportionnelle à [A]
Ordre 2
V = K · [A]²
Vitesse proportionnelle au carré de [A]
En cinétique enzymatique, on observe une transition entre ordre 1 et ordre 0 selon la concentration en substrat — c’est l’une des caractéristiques les plus importantes du comportement enzymatique.
III. Le Modèle de Michaelis-Menten : La Théorie Fondamentale
En 1913, Leonor Michaelis et Maud Menten publient une théorie révolutionnaire : l’enzyme (E) et son substrat (S) se combinent de façon réversible pour former un complexe intermédiaire ES. Ce complexe se dissocie ensuite pour libérer le produit (P) et régénérer l’enzyme libre :
E + S ⇌ ES → E + P
k₁ k₋₁ k₂
L’idée centrale est que la formation du complexe ES est obligatoire pour que la catalyse ait lieu. L’enzyme n’est pas consommée ; elle retrouve sa forme initiale après chaque cycle.
3.2 L’Hypothèse de la Vitesse Initiale
Pour simplifier l’analyse, on mesure la vitesse au tout début de la réaction, alors que :
La concentration en produit [P] est encore quasi nulle
La réaction inverse (E + P → ES) est donc négligeable
Les concentrations en E et S restent pratiquement constantes
Cette condition expérimentale rend les équations mathématiques beaucoup plus maniables.
3.3 L’Hypothèse de l’État Stationnaire (Briggs-Haldane, 1925)
Briggs et Haldane ont affiné le modèle en 1925 avec un postulat plus général : la concentration en complexe [ES] reste constante pendant la phase initiale de la réaction, car sa vitesse de formation est égale à sa vitesse de disparition.
Vformation ES = Vdisparition ES
k₁ · [E] · [S] = (k₋₁ + k₂) · [ES]
Cette hypothèse d’état stationnaire est aujourd’hui universellement utilisée en enzymologie.
IV. L’Équation de Michaelis-Menten et l’Interprétation de la Courbe
4.1 L’Équation
En combinant la conservation de la concentration totale d’enzyme ([E]T = [E] + [ES]) avec l’hypothèse de l’état stationnaire, on obtient l’équation de Michaelis-Menten :
V = (Vmax · [S]) / (KM + [S])
C’est une fonction hyperbolique de la concentration en substrat, avec deux paramètres caractéristiques : Vmax et KM.
4.2 Interprétation de la Courbe
La courbe V = f([S]) présente trois régimes distincts :
Faible [S] : V est proportionnelle à [S] → comportement de premier ordre. L’enzyme a des “places libres” ; plus on ajoute de substrat, plus la réaction est rapide.
[S] intermédiaire : la relation n’est plus linéaire → ordre mixte. L’enzyme commence à être saturée.
[S] élevée : V atteint un plateau → ordre zéro, V ≈ Vmax. Toutes les molécules d’enzyme sont occupées. Ajouter du substrat supplémentaire ne change plus rien. C’est le phénomène de saturation enzymatique, propre à la cinétique enzymatique.
V. Signification et Importance des Constantes Cinétiques
5.1 La Vitesse Maximale (Vmax)
La Vmax correspond à la vitesse de la réaction lorsque toutes les molécules d’enzyme sont saturées par le substrat, c’est-à-dire quand [ES] = [E]T :
Vmax = k₂ · [E]T
La Vmax est directement proportionnelle à la quantité totale d’enzyme dans le milieu. C’est ce principe qui est exploité en biologie clinique pour doser indirectement la concentration d’une enzyme dans un échantillon biologique (sérum, tissu).
Exemple clinique : la mesure des transaminases (ASAT, ALAT) dans le sérum sanguin, via leur Vmax, permet de détecter des lésions hépatiques comme l’hépatite ou la cirrhose.
5.2 La Constante de Michaelis KM
La KM est la concentration en substrat pour laquelle la vitesse est égale à la moitié de la Vmax (V = Vmax/2). Elle est définie par :
KM = (k₋₁ + k₂) / k₁
La KM mesure l’affinité de l’enzyme pour son substrat :
KM faible → haute affinité (l’enzyme se sature à faible concentration de substrat)
KM élevée → faible affinité (il faut beaucoup de substrat pour saturer l’enzyme)
La KM est une constante caractéristique du couple enzyme-substrat. Elle varie avec le pH, la température et la nature chimique du substrat.
Exemple concret : l’hexokinase cérébrale possède un KM pour le glucose de ~0,1 mM, tandis que la glucokinase hépatique a un KM d’environ 10 mM. Cette différence permet au cerveau de capter en priorité le glucose sanguin, même en faibles concentrations.
5.3 La Constante Catalytique kcat (Turnover Number)
La kcat, aussi appelée nombre de renouvellement, représente le nombre de molécules de substrat converties en produit par molécule d’enzyme et par unité de temps, lorsque l’enzyme est saturée :
kcat = Vmax / [E]T (unité : s⁻¹)
C’est une mesure directe de la performance maximale de l’enzyme. Quelques valeurs remarquables :
Enzyme
kcat (s⁻¹)
Anhydrase carbonique
~10⁶
Catalase
~4 × 10⁷
Chymotrypsine
~100
Lysozyme
~0,5
5.4 La Constante de Spécificité kcat/KM
Dans les conditions cellulaires réelles, la concentration en substrat [S] est rarement saturante. Elle se situe généralement entre 0,01 et 1 fois la KM. Dans ces conditions, kcat seule ne suffit pas pour évaluer l’efficacité d’une enzyme.
La constante de spécificité kcat/KM est la grandeur la plus pertinente pour mesurer l’efficacité catalytique en conditions physiologiques :
kcat/KM = constante de vitesse pour l'interaction E + S → produit
Elle prend en compte à la fois la vitesse de catalyse (kcat) et la force d’interaction avec le substrat (KM).
Limite théorique et perfection catalytique : la valeur maximale de kcat/KM est imposée par la vitesse de diffusion des molécules en solution, soit environ 10⁸ à 10⁹ M⁻¹·s⁻¹. Les enzymes qui atteignent cette valeur sont dites “catalytiquement parfaites” — elles transforment le substrat pratiquement à chaque collision.
Exemple : la catalase et l’anhydrase carbonique sont considérées comme des enzymes parfaites avec des kcat/KM proches de 10⁸–10⁹ M⁻¹·s⁻¹.
Cette constante permet également de comparer la spécificité d’une enzyme pour différents substrats : l’enzyme préférera toujours le substrat ayant la valeur de kcat/KM la plus élevée.
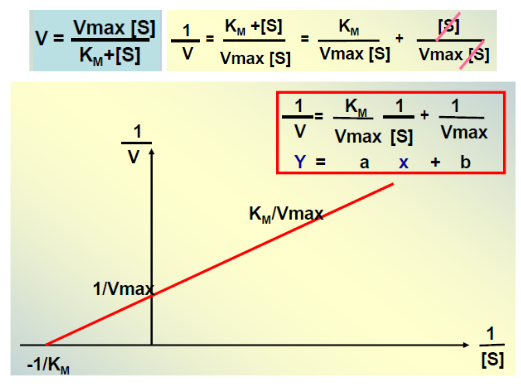
VI. Détermination Graphique de Vmax et KM : La Représentation de Lineweaver-Burk
Figure 3 : Graphique de Lineweaver-Burk — la linéarisation permet de déterminer Vmax et KM avec précision.
6.1 Pourquoi Linéariser ?
La courbe hyperbolique de Michaelis-Menten ne permet pas une détermination graphique précise de Vmax et KM, car l’hyperbole atteint son asymptote très progressivement. On procède donc à une transformation mathématique pour obtenir une droite plus facile à exploiter.
6.2 La Méthode des Doubles Inverses (Lineweaver & Burk)
En prenant l’inverse des deux membres de l’équation de Michaelis-Menten, on obtient :
1/V = (KM/Vmax) · (1/[S]) + 1/Vmax
C’est l’équation d’une droite y = ax + b où :
La pente = KM / Vmax
L’ordonnée à l’origine = 1/Vmax → on en déduit Vmax
L’abscisse à l’origine = -1/KM → on en déduit KM
Avantages : détermination précise des paramètres cinétiques par régression linéaire.
Limites : les points aux faibles [S] (grandes valeurs de 1/[S]) ont un poids excessif et peuvent fausser la droite. D’autres représentations linéarisées existent (Eadie-Hofstee, Hanes-Woolf) pour pallier ces inconvénients.
Conclusion et Points Clés à Retenir
La cinétique enzymatique est bien plus qu’un exercice mathématique : c’est un outil conceptuel puissant pour comprendre comment les enzymes régulent le métabolisme cellulaire et comment les médicaments peuvent moduler leur activité.
Les points essentiels de cette première partie :
La cinétique enzymatique étudie la vitesse des réactions catalysées par les enzymes en fonction de divers paramètres
Le modèle de Michaelis-Menten (1913), affiné par Briggs-Haldane (1925), repose sur la formation d’un complexe intermédiaire ES
La courbe V = f([S]) est hyperbolique et montre trois régimes : ordre 1, mixte, puis ordre 0 (saturation)
Vmax = vitesse maximale atteinte à saturation ; KM = mesure de l’affinité enzyme-substrat
La représentation de Lineweaver-Burk permet la détermination graphique précise de KM et Vmax
Dans la Partie 2, nous aborderons les inhibiteurs enzymatiques (compétitifs, non compétitifs, incompétitifs) et l’effet des facteurs physico-chimiques (pH, température) sur l’activité enzymatique.
Mots-clés : cinétique enzymatique, modèle de Michaelis-Menten, constante KM, Vmax, kcat, turnover number, Lineweaver-Burk, vitesse de réaction enzymatique, état stationnaire, complexe enzyme-substrat, biochimie, pharmacologie enzymatique

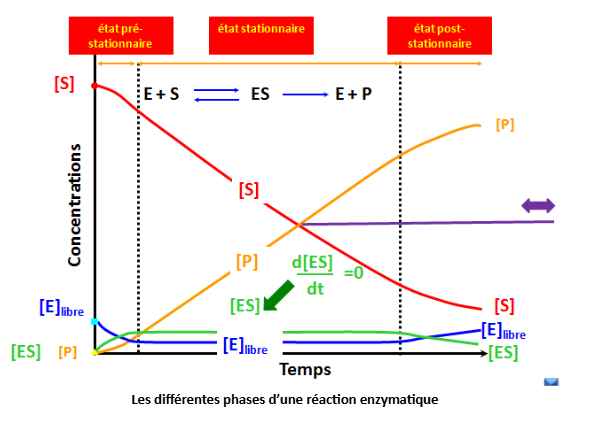
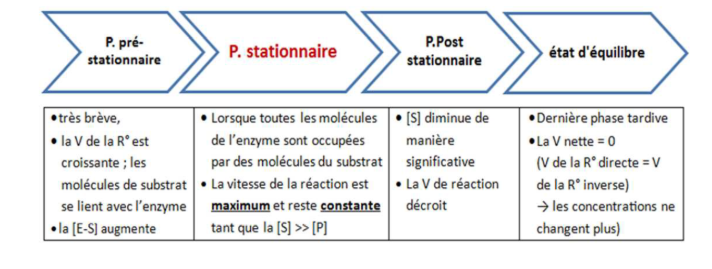
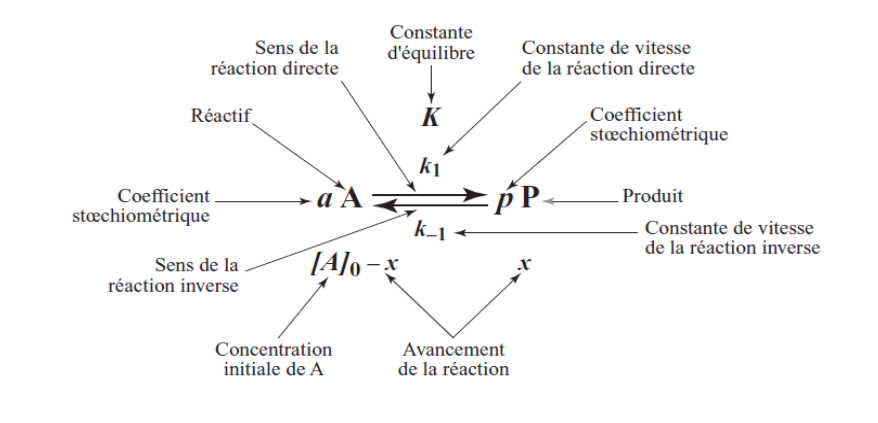
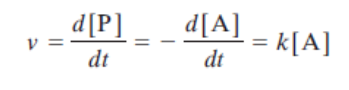
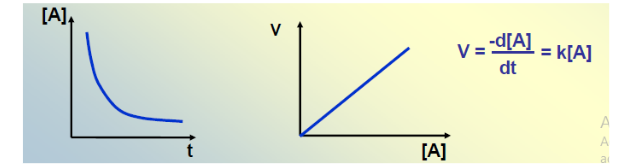
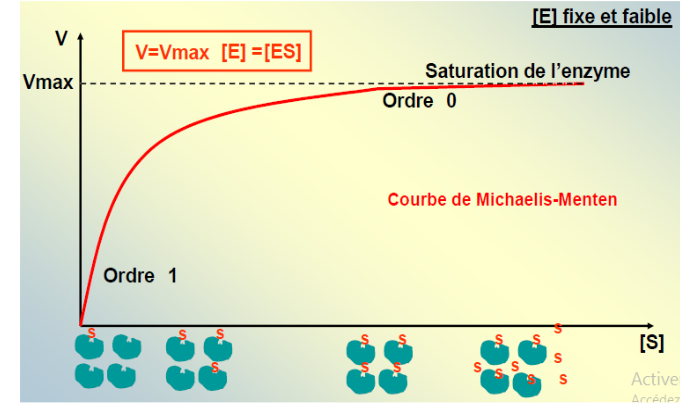
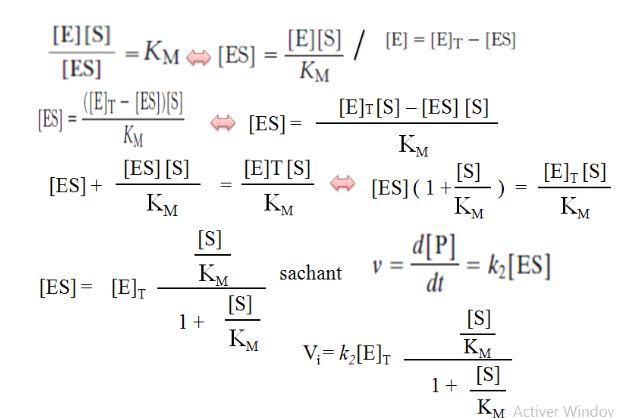
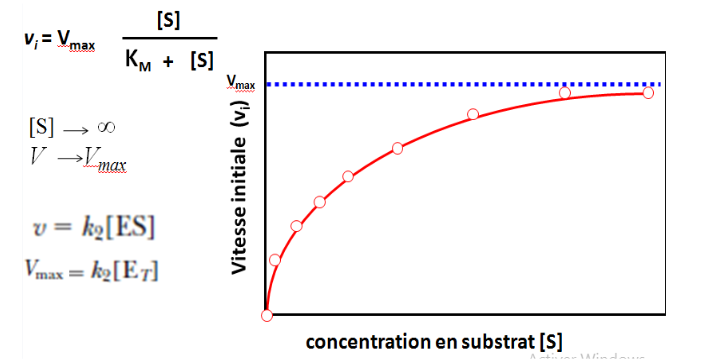
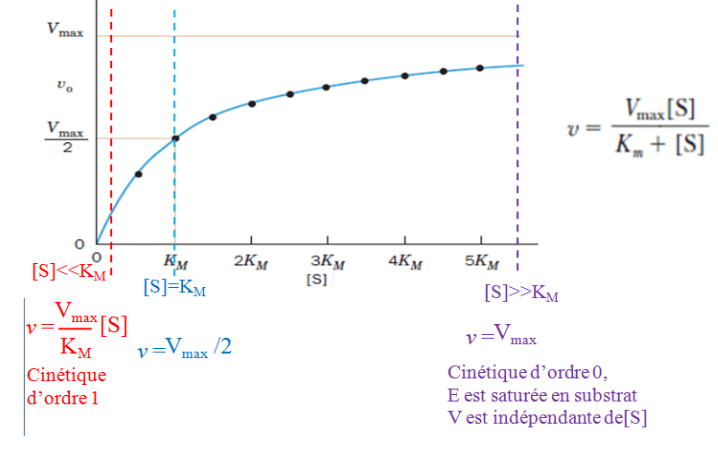
 Figure 3 : Graphique de Lineweaver-Burk — la linéarisation permet de déterminer Vmax et KM avec précision.
Figure 3 : Graphique de Lineweaver-Burk — la linéarisation permet de déterminer Vmax et KM avec précision.